Archaeopteryx, the earliest known bird
History of Archaeopteryx
The fossil bird Archaeopteryx is one of the most famous of all extinct organisms. Archaeopteryx fossils are known only from 150‐million‐year‐old limestone deposits in southern Germany. The species name Archaeopteryx lithographica derives from the fact that most specimens come from rock quarries that were mined for use in lithography, a printing process. Fossils of Archaeopteryx and thousands of other Jurassic Period organisms were found when quarry workers split slabs of this stone to make lithographic plates.
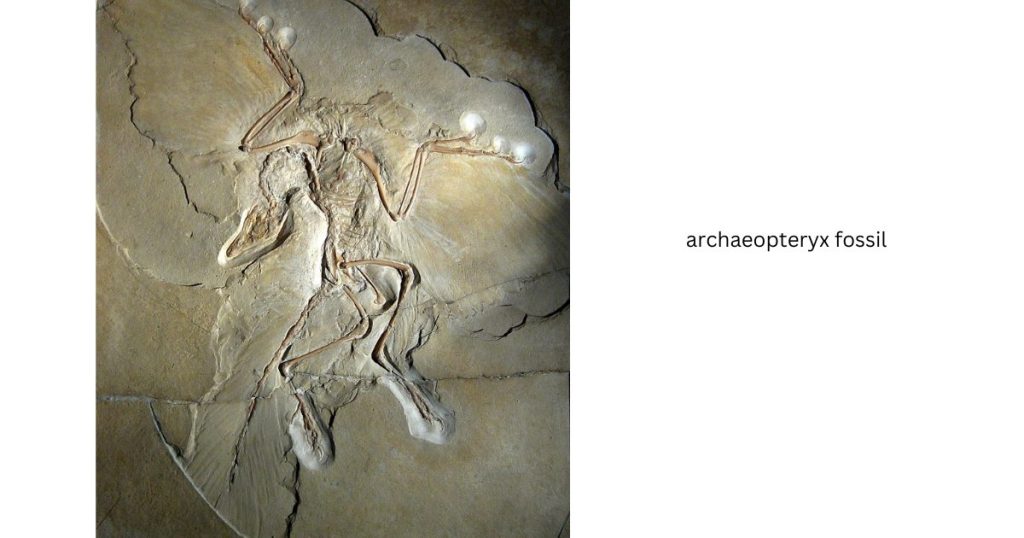
Earliest Archaeopteryx specimens
Several of the earliest Archaeopteryx specimens found in the 1860s and 1870s were almost complete skeletons, with clear impressions of feathers surrounding the arms and body. One of the most spectacular of these fossils is now in the natural history museum at Humboldt University in Berlin, Germany; it accordingly is referred to as the “Berlin specimen”. Another famous example known as the “London specimen” is in the British Museum. The limestone sediments from which all Archaeopteryx fossils have been found were formed in a shallow, tropical sea dotted with islands. Other flying vertebrates coexisted there with Archaeopteryx, including a variety of flying reptiles known as pterosaurs. Pterosaur fossils are much more common in these deposits than Archaeopteryx fossils.

Modern bird traits in Archaeopteryx fossil
The early Archaeopteryx fossils were and remain sensational finds because they reveal an organism with a mosaic of traits showing linkages to both theropod dinosaurs and modern birds.

Bird‐like traits in Archaeopteryx include the structure of its plumage, its relatively large brain, and many details of its skeletal anatomy. Features that link it to its non‐avian theropod relatives include its many small teeth, a long tail with a bony midline, claws on the ends of its three finger bones in the wings, and extraskeletal details.
Does Archaeopteryx fly?
There is still great debate about the flying ability and behavior of Archaeopteryx. The largest specimen of this species had a wingspan of about half a meter, and various features of its wing and feather anatomy suggest that it could fly, but it is not clear how well. The claws on its wings have featured prominently in reconstructions of Archaeopteryx as a climbing, arboreal specialist, but other anatomical features suggest that it may have spent much of its time on the ground.
Is Archaeopteryx a dinosaur or a bird?
Despite the clear affinities of Archaeopteryx to both modern birds and extinct groups of dinosaurs, we now understand that Archaeopteryx is not a direct missing link between these groups. This is because Archaeopteryx is not a direct ancestor of modern birds, but instead a representative of an extinct evolutionary offshoot of early birds, a side branch of the large diversity of early birds that originated earlier in the Jurassic Period.
A different and currently unknown early bird present at this time was more likely the direct ancestor of all modern birds. Nonetheless, Archaeopteryx is generally considered to be the oldest known organism that is a bird. New discoveries of contemporaneous theropods with avian attributes highlight the great diversity of such lineages in the late Jurassic fauna. As in all situations where diverse lineages occur at a time of great evolutionary transition, and because all fossils are inherently incomplete, it can be challenging to distinguish the early birds from non‐avian theropods with many similar traits.
Other early birds from the Mesozoic
Animals that were unquestionably birds exploded in diversity during the last part of the Mesozoic Era, in the Cretaceous Period from 66 to 145 million years ago. However, most of the fossil birds from this time are not the direct ancestors of the modern birds today; instead, they are offshoots of the avian evolutionary tree that left no direct descendants. Of the dozens of avian groups present in the Cretaceous that are now extinct, four merit special recognition.
Confuciusornis
Discovered in 1993, the crow‐sized Confuciusornis is now one of the best-understood Mesozoic birds due to the exquisite level of anatomical detail preserved in some fossils.
Although they are known from the Liaoning deposits found in China that date to about 120–135 million years ago, hundreds of Confuciusornis specimens have been studied by avian paleontologists; unfortunately, hundreds, or perhaps thousands, of additional specimens have been often sold illegally to collectors and are not available for research. The skeletal anatomy of Confuciusornis has many avian characters but lacks adaptations seen in modern birds,
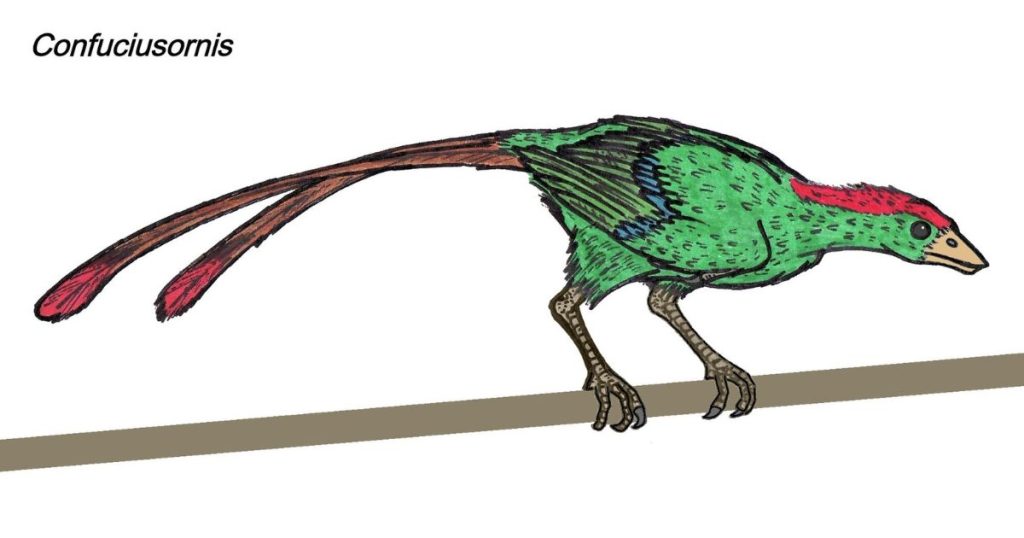
such as an extensive keel on the sternum. Its wings were long and pointed, and its wing feathers were highly asymmetrical, as seen in modern flying birds. Instead of the long, bony tail seen in earlier birds such as Archaeopteryx, Confuciusornis had a pygostyle, the knot of fused vertebrae that support the tail feathers in modern birds. Although Confuciusornis did not have the full, fan‐shaped tail seen in most living birds, many specimens preserve two dramatically long tail plumes. These feathers are reminiscent of sexually selected ornaments seen in many groups of living birds and suggest that female birds of various species have been selecting mates with long tails for at least 140 million years.
Hesperornis
Some groups of Mesozoic birds, including Hesperornis and its relatives in the order Hesperornithiformes, have been known to paleontologists for much longer than Confuciusornis. Hesperornis fossils were discovered in the western part of North America in the 1880s. These birds were all highly specialized swimmers with long, torpedo‐shaped bodies.

Their reduced wings indicate that, like penguins, they had lost the ability to fly. With large, strong leg muscles and lobed feet, they could move swiftly underwater to catch fish in their long, toothed jaws. More fragmentary remains of related species indicate that Hesperornis is but one representative of a larger radiation of birds with similar features that went extinct at the close of the Cretaceous Period.
Enantiornithes
Fossils of one of the most successful and diverse groups of Mesozoic birds, the Enantiornithes, are found on all modern continents except Antarctica. The diversity of enantiornithine birds may have rivaled the varied foraging and morphologies specializations seen in modern songbirds.

Enantiornithes were similar in body size to today’s songbirds, and their skeletal details suggest that many were strong fliers. Although their finger bones had claws, the general structure of enantiornithine wings shares many features including the presence of an alula with modern birds. Considered a group, the Enantiornithes had diverse ecologies, including both aquatic and terrestrial lineages. Fossil evidence from tiny droplets of amber indicates that at least one of these species had an unusual diet: fossilized tree resin seen in the bird’s digestive tract suggests that this enantiornithine species fed literally on tree sap.
Ichthyornis
Ichthyornis is another well‐known bird from the Mesozoic. Compared with Enantiornithes, Hesperornis, and Confuciusornis, Ichthyornis is probably more closely related to the lineage that has given rise to modern birds, although it too is not their direct ancestor. Ichthyornis was a fish‐eating, strong flier equivalent of modern seabirds, and was the size of a modern tern. Its wing structure and breastbone with a large keel were similar to modern birds. Many Ichthyornis specimens were collected in western North America in the late 1880s, and, along with Hesperornis, the discoveries of early birds with teeth became well known. In 1880, Charles Darwin wrote a letter to an American paleontologist noting that these archaic birds provided some of “the best support to the theory of evolution” since his publication of his Origin of Species.

Despite the recent dramatic discoveries of fossil lineages of Mesozoic birds, we only have fragmentary evidence for the full diversity of early birds and other bird‐like dinosaurs from this time period. Only a single lineage from the entire radiation survived until the current time to become the lineage of modern birds.

Mass extinction and the origin of modern birds
There is ongoing debate as to when the lineages of modern birds living today originated and started to diversify from one another. The devastating mass extinction event that ended the Cretaceous Period 65.5 million years ago is most famous for causing the extinction of all dinosaurs except for birds. This end‐Cretaceous event also caused the demise of many other groups of terrestrial and marine organisms, and it may have sparked the diversification of many of the bird lineages that still exist today.
Does climate shift caused the mass extinction?
Although the causes of this mass extinction are long obscure, evidence suggests that the impact of a huge asteroid, which produced the Chicxulub crater near Mexico’s Yucatan Peninsula,
resulted in sudden shifts in climate worldwide. However, independent events happening at the same time,
such as sharp rises in sea levels and volcanic activity, have contributed to the mass extinction.
Did the asteroid hit cause this?
Fossil deposits from just before the asteroid impact are particularly important for pinpointing whether the extinction was rapid as would be expected if it resulted in primarily from the asteroid impact or more gradual, as would be expected if there were multiple factors.
The few deposits of avian fossils found just before 65.5 million years ago suggest that the asteroid impact was devastating to the birds present in the late Mesozoic. For example, analyses of avian bones from sites in North America and Madagascar have revealed a wide diversity of Mesozoic birds, including representatives of the Hesperornithiformes Enantiornithes, and Ichthyornis, that persisted until the very end of this period but not beyond. In contrast, these surveys did not identify any bones that could be assigned to orders of birds still living today, suggesting that the diversification of most modern groups of birds started after the mass extinction event.
In fact, there is only one known fossil from the entire Cretaceous that unambiguously falls in a group of still-living birds: the fossil duck relative Vegavis. Many other Cretaceous fossils from sites around the world have been linked tentatively to groups of living birds, but these associations usually are uncertain and often based on single bones. In contrast, the Vegavis fossil specimen is a nearly complete skeleton, except for the skull.
The Vegavis fossil was found in Antarctica in 1992 but was not recognized as a waterfowl until detailed analyses were made a decade later. The presence of a duck relative in the late Cretaceous has important implications because we know from other phylogenetic evidence that geese, ducks, and their relatives in the Anseriformes split from the group that includes living grouse, chickens, and pheasants (order Galliformes) after an earlier split that divided the paleognaths (kiwis, ostriches, emus, tinamous, and their relatives) and all other living birds. The fossil Vegavis shows that these two earlier splits must predate the end of the Cretaceous, despite the existing lack of fossil evidence for these modern lineages during the Mesozoic.
After effects of mass extinction:
Fossils representing many modern orders are seen to be found in subsequent deposits from the early Tertiary Period, after the mass extinction. However, several archaic bird lineages that were common in the late Cretaceous are absent in the Tertiary. This quick turnover pattern in the avian fossil record also suggests the fact that almost all bird lineages were wiped out completely during the mass extinction at the junction between the Tertiary and Cretaceous. Under this line of reasoning, the few groups of birds survived and then rapidly diversified in the very early part of the Tertiary as the environment became more hospitable, refilling many avian ecological niches opened up by the mass extinction. This period of very rapid radiation in the early Tertiary gave rise to many modern families and orders.
Molecular clock method
In contrast to the later, Tertiary radiation implied by the absence of most modern birds from the earlier fossil record, studies conducted on genetic comparisons consistently point to a much earlier origin of modern birds. These DNA dates are based on evolutionary trees calibrated using fossils from deposits of known age. Dates from a few well-documented fossils can help calibrate the timescale for more extensive phylogenies that are mostly based on DNA comparisons among still-living species. This molecular clock method is useful for back‐calculating the point in the past when two bird lineages diverged from one another.
Different mutations accumulate over time in each descendant lineage once they separate from their common ancestor; therefore, if you know the overall average rate of mutational change, you can use the amount of DNA difference between two living species to calculate the time when their ancestors separated. In practice, this process is not very accurate, in part because different parts of the genome have different average mutation rates. After all, well‐dated fossils are absent for many groups of living birds and other technical reasons. Many of these procedural issues can be addressed via statistical analyses that take these underlying sources of variation into account to provide an idea of the most likely time for each evolutionary separation, rather than pinpointing one exact age.
Using the molecular clock approach to date the origins of groups of organisms was initially controversial when the method was introduced several decades ago, but today it is widely applied and broadly accepted among evolutionary biologists.
Final finding
Controversies involving the outcomes of analyzing molecular clocks still exist, however, including the general discrepancy between the DNA and fossil dates for the origins of most groups of modern birds. For reasons that remain uncertain, dates estimated using molecular tools are almost always much earlier than the corresponding earliest fossils known for those groups. Even the most advanced DNA‐based estimates suggest that the earliest split within the evolutionary tree of living birds the split between the paleognaths and all other living birds occurred in the earlier part of the Cretaceous, at roughly 130 million years ago. Some studies further suggest a Cretaceous origin for many living orders, yet the earliest equivalent fossils found of nearly all of these groups are from much later in the Tertiary Period.
When both the fossil and genetic evidence are considered, it seems clear that at least a few lineages of modern birds arose before the mass extinction at the Cretaceous-Tertiary boundary. The extent of the Cretaceous radiation of modern birds remains uncertain, but future fossil discoveries may provide direct evidence for the existence of the ghost lineages in the Cretaceous whose existence currently is inferred from phylogenies but which are missing physically from the known fossil record.
Additional genome-wide DNA data, analyzed in concert with improved molecular clock analyses and calibration, alternatively might show that most of the diversification in modern birds occurred in the Tertiary, as these lineages expanded into niches opened in the previous mass extinction. Reconciling these two possibilities remains a significant challenge in avian systematics.